


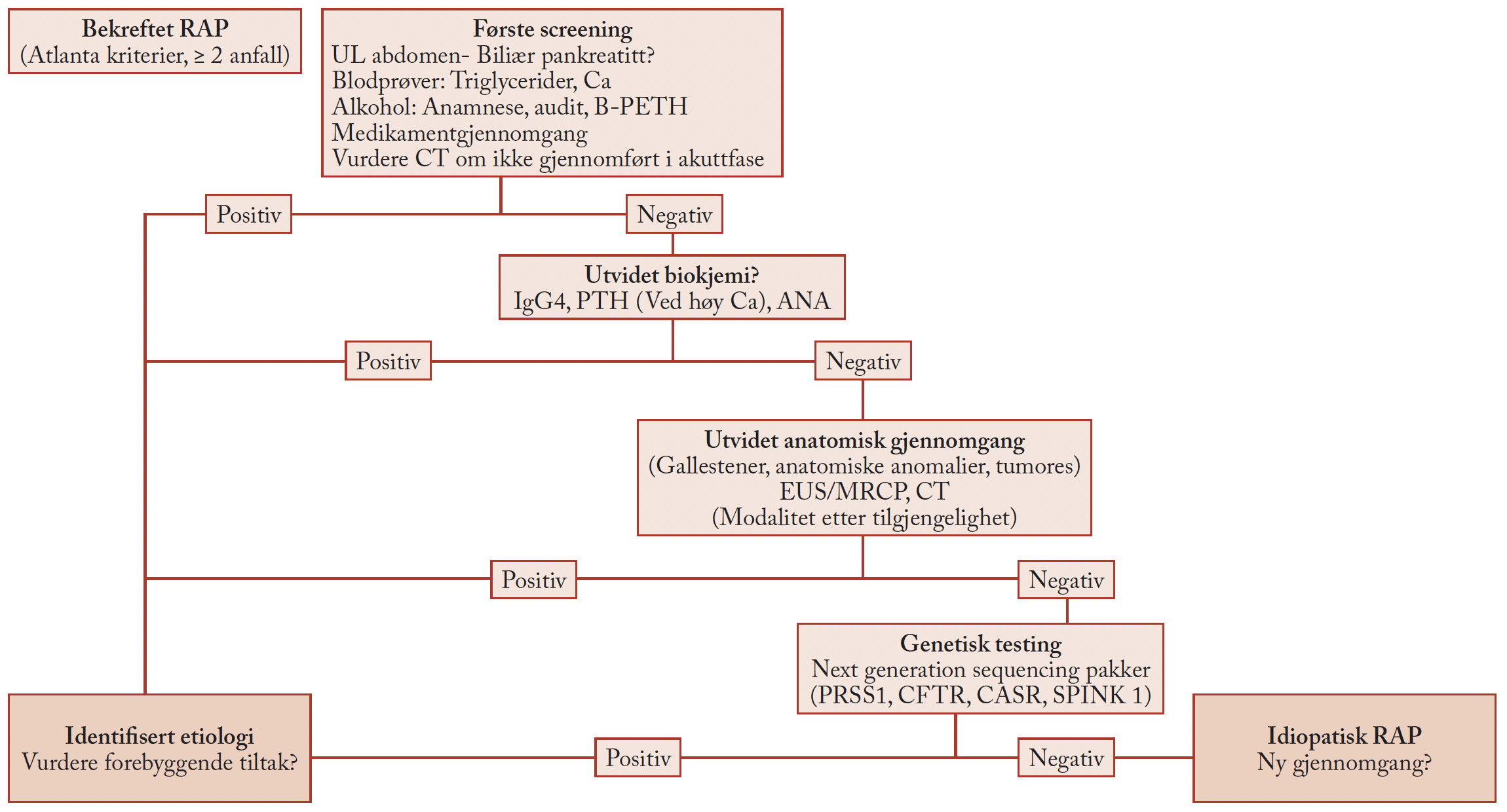
Tone Møller Tannæs.
Tekst: Postdoc Tone Møller Tannæs og stipendiat Hilde Synnøve Vollan.
Avdeling for Klinisk Molekylærbiologi, Akershus universitetssykehus og Universitetet i Oslo
Hva som gjør at noen lever hele livet med en Helicobacter uten problemer, mens det hos andre utvikles sykdom er ikke klarlagt. Mange virulensfaktorer er studert, men ingen av disse kan gi en absolutt forklaring. Noe av ‘problemet’ er at det er samme bakteriestamme man bærer på hele livet – den byttes ikke ut med en mer aggressiv slektning. Det er dette vår forskningsgruppe ved Avdeling for klinisk Molekylærbiologi ved Akershus universitetssykehus jobber med.

Hilde Synnøve Vollan.
H. pyloris genomsekvens er sterkt konservert, og vi finner hovedsakelig de samme genene hos ulike bakterieisolater. Allikevel har H. pylori mye større genetisk diversitet enn det som er observert hos andre bakteriearter. Denne enorme diversiteten skyldes blant annet at bakterien naturlig kompetent og kan ta opp DNA fra omgivelsene. H. pylori har dermed en fascinerende evne til kontrollert variasjon av sitt genmateriale. Det ustabile genomet benytter bakterien for å kunne tilpasse seg til ulike miljømessige situasjoner. I magen er det blant annet store døgnvariasjoner i pH, verten vil endre seg gjennom titalls år med kolonisering og på et tidspunkt må dessuten bakterien ha oppholdt seg utenfor kroppen på vei til en ny kropp.

Figur 1. Mekanismen bak fasevariasjon.
En av de genetiske mekanismene som H. pylori benytter for å variere genuttrykket sitt er fasevariasjon (Fig. 1). Figuren viser at polymerasen ’glipper’ under DNA-syntesen (DNA slippage) og genet mister eller får tilført ekstra nukleotider i den nylagde bakterien. Konsekvensene av fasevariasjonen er at genene kan variere mellom å kode for funksjonelle og ikke-funksjonelle produkter.

Figur 2. Effekten av fasevariasjon i pldA-genet.
Vår forskningsgruppe har funnet at pldA, genet som koder for fosfolipase A (OMPLA) hos H. pylori, er fasevariabelt (1). OMPLA er et enzym som spalter bakteriens fosfolipider til lysofosfolipider. Fasevariasjonen gir bakterien en mulighet til å skru OMPLA-aktiviteten PÅ eller AV. Bakterievariantene med en aktiv OMPLA karakteriseres ved at de har en cellevegg med svært høyt innhold av lysofosfolipider. Denne celleveggen vil være mindre tett og ha større tendens til å slippe ut proteiner (Fig. 2) (2). Vi har vist at denne fasevariasjonen kan være assosiert med bakteriens sykdomsfremkallende evne (3).

Figur 3. Modell av H. pylori OMPLA.
For å forstå denne prosessen med hensyn på proteinets funksjon i forhold til sykdomsutvikling, er det nødvendig å kjenne OMPLAs tredimensjonale struktur. Vi har derfor konstruert en proteinstrukturmodell. Vi har sammenlignet DNA-sekvenser av OMPLA-lignende proteiner fra ca 1000 ulike bakteriearter. Med dette som bakgrunn er det etablert en simulert proteinmodell av H. pyloris OMPLA (Fig. 3) (4).

Figur 4. H. pyloris OMPLA kan være involvert i protontransport.
Den foreløpige modellstrukturen legges til grunn for en ny modell for hvordan H. pylori interagerer med sin vert. Modellstrukturen gir dessuten grunnlag for å danne hypoteser om nye funksjoner knyttet til dette yttermembranproteinet. Vi mistenker utfra datasimuleringer at OMPLA kan danne en pore som er koblet til protontransport (Fig. 4). Det pågår samtidige studier i laboratoriet for å validere de funn som kommer ut av modellen. Vi ser for eksempel at alle endringer i OMPLA som ødelegger fosfolipaseaktiviteten, gjør at bakterien mister evnen til å overleve ved lav pH. En observasjon som kan tyde på at OMPLA er involvert i protontransport.
For å validere den simulerte OMPLA-strukturen forsøker vi å krystallisere OMPLA på laboratoriet. Vi har uttrykt H. pyloris OMPLA i E. coli, og vi jobber nå med å få foldet det denaturerte proteinet korrekt. Når vi har oppnådd et foldet, aktivt protein vil røntgenkrystallografiske analyser vise oss hvordan proteinstrukturen ser ut i virkeligheten. Krystallstrukturen vil forhåpentligvis blant annet gi oss en forklaring på hvorfor OMPLA er nødvendig for at H. pylori skal overleve ved lav pH.
Referanser
- Tannaes, T., N. Dekker, et al. (2001). ”Phase variation in the Helicobacter pylori phospholipase A gene and its role in acid adaptation.” Infection and Immunity 69(12): 7334-7340.
- Bukholm, G., T. Tannaes, et al. (1997). ”Colony variation of Helicobacter pylori: Pathogenic potential is correlated to cell wall lipid composition.” Scandinavian Journal of Gastroenterology 32(5): 445-454.
- Tannaes, T., I. K. Bukholm, et al. (2005). ”High relative content of lysophospholipids of Helicobacter pylori mediates increased risk for ulcer disease.” Fems Immunology and Medical Microbiology 44(1): 17-23.
- Vollan, H. S., G. Portella, et al. (2008). ”A homology model of H-pylori OMPLA.” Helicobacter 13(5): 423-423.
Forfatter
Related Posts
Neste kurs/møte
